Woda wraz z solami mineralnymi jest pobierana przez rosliny z gleby za pomocą korzeni. Następnie jest transportowana do łodyg i liści za pośrednictwem tkanki przewodzącej - drewna. Woda jest wykorzystywana w procesie fotosyntezy, który zachodzi głównie w liściach. Związki ograniczne powstałe w wyniku tego procesu są transportowane do pozostałych organów rośliny pośrednictwem łyka.
Woda jest składnikiem każdej żywej komórki. Jej zawartość w tkankach waha się od 10-15% w nasionach do ponad 90% w płatkach kwiatów. Życie bez wody jest niemożliwe, ponieważ pełni ona wiele istotnych funkcji m.in.:
Korzenie pobierają składniki mineralne najczęściej w postaci jonów. Głównym ich źródłem dla roślin jest roztwór glebowy, czyli zawarta w glebie woda z rozpuszczonymi w niej solami mineralnymi. Roślina czerpie z gleby tylko potrzebne jej jony. Dzieje się tak nawet wówczas gdy ich stężenie w roztworze glebowym jest mniejsze niż we włośnikach, czyli wbrew różnicy stężeń. Jest to możliwe dzięki znajdującym się w błonach komórkowych włośników wyspecjalizowanym białkom nośnikowym. Umożliwiają one przemieszczanie sie jonów na drodze transportu aktywnego, a więc z udziałem energii.

Rośliny pobierają wodę przez korzenie. Najintensywniaj odbywa sie to w ich strefie włośnikowej. Główną rolę w procesie pobierania wody przez komórki roślin odgrywają zjawiska osmotyczne.Cząsteczki wody są pobierana i transportowane przez półprzepuszczalne błony komórkowe zgodnie z różnicą stężeń, a tym samym - z różnicą potencjału wody. Stanowi on miarę zdolności komórki do pobierania lub oddawania wody na zasadzie osmozy. Przyjmuje się że potencjał czystej wody Ψ (Psi) wynosi 0 jednostek, przy czym jednostką potencjału wody jest pascal (Pa). Rozpuszczenie w wodzie substancji powoduje spadek potencjału wody, ma on wówczas wartość ujemną.
Potencjał wody w komórce zależy od ciśnienia hydrostatycznego oraz ciśnienia osomotycznego. Ciśnienie hydrostatyczne, zwane inaczej turgorowym, to nacisk, jaki wywiera cytoplazma otaczająca jądro komórkowe na ścianę komórkową. Spadek ciśnienia hydrostatycznego jest spowodowany utratą wody i objawia się więdnięciem organów rosliny. Ciśnienie osmotyczne to siła, z jaką cząsteczki rozpuszczonych substancji przyciągają cząsteczki rozpuszczalnika (wody) przez błonę półprzepuszczalną. Zależy ono od stężeń roztworów po obu stronach błony. Im wieksze stężenie roztworu w komórce, tym większe jest jego ciśnienie osomotyczne, a w rezultacie tempo osmozy, czyli siła z jaką komórka pobiera wodę. Zależność między potencjalem wody, cośnieniem tugorowym, a ciśnieniem osmotycznym wyraża poniższe równanie:
ΨK = P - π , gdzie
ΨK - potencjał wody w komórce
P - ciśnienie tugorowe
π - ciśnienie osmotyczne roztworu
Woda przepływa przez błonę komórkową zawsze z roztworu o wyższym potencjale wody do roztworu o niższym potencjale wody. Roztwór o wyższym potencjale wody ma wyższe stężenie wody, a niższe stężenie substancji rozpuszczonej niż roztwór o niższym potencjale wody. Najwyższą wartość ma potencjał wody w glebie, a najniższą - w atmosferze. To powoduje że woda stale wnika z gleby do korzeni, przepływa przez łodygi i liście, a nastepnie przedostaje sie do atmpsfery w postaci pary wodnej.
Pobieranie i transport wody w roslinie zachodzą przede wszystkim biernie, bez nakładów energii. Jest to mozliwe dzięki transpiracji - parowaniu wody z nadziemnych części rosliny, które odbywa się głównie przez aparaty szparkowe i przedchlinki. Ciągły ubytek wody umożliwia utrzymanie różnicy jej potencjału, a w konsekwencji - stały przepływ wody przez roślinę. Transpiracja powoduje wytworzenie w naczyniach podciśnienia hydrostatycznego, które sięga od liści, przez łodygę aż do korzeni i działa jak pompa ssąca. Proces ten przypomina zasysanie płynu przez słomkę. Warunkiem transportu wody w naczyniach jest istnienie nieprzerwanego słupa wody. Utrzymanie go jest możliwe dzieki:
W wypadku braku lub ograniczenia transpiracji znaczenia nabiera mechanizm czynny pobierania wody, który zazwyczaj wspomaga jedynie transport wody i soli mineralnych. Wemaga on dopływu energoo uzyskiwanej w procesie oddychania. Mechanizm ten polega na aktywnym pobieraniu z gleby przez komórki korzenia soli mineralnych, które są następnie transportowane do elementów przewodzących - drewna.
napływ wody hest przyczyną wytworzenia w elementach przewodzących drewna ciśnienia, tzw. parcia korzeniowego, które działa jak pompa tłocząca wodę w górę. Przejwaem parcia korzeniowego jest gutacja, czyli wydzielanie kropli płynu na brzegach i wierzchołkach liści przez specjalne otwory - hydatody oraz tzw.

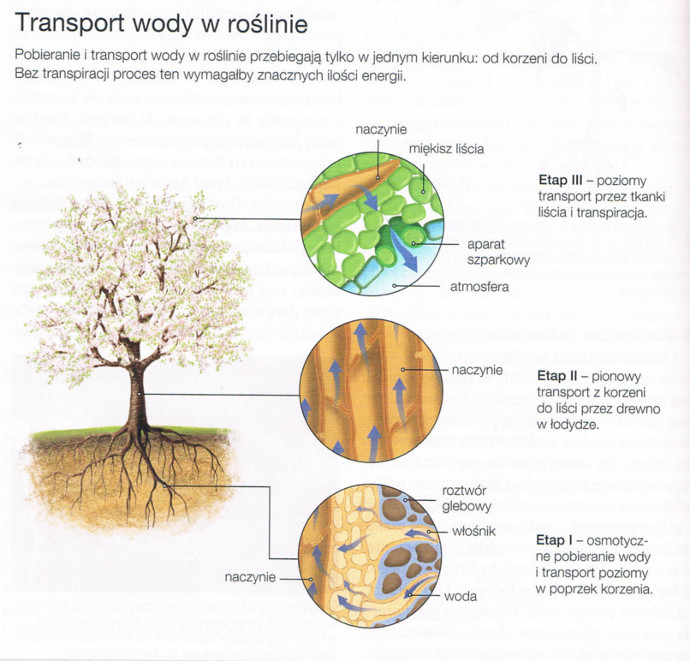
Transport wody i soli mineralnych w układzie gleba - roślina - powietrze zachodzi w trzech etapach:
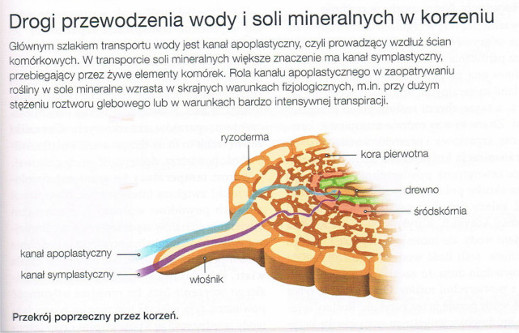
Woda i sole mineralne pobrane przez włośniki są transportowane przez komórki kory pierowtnej do walca osiowego, a stamtąd do wnętrza elementów przewodzących. Transport w poprzek tkanek korzenia odbywa się dwoma kanałami:
Woda i sole mineralne są przenoszone z korzeni do liści przez elementy przewodzące w łodydze. Transport pionowy wody i soli mineralnych może osiągaś prędkońć od kilku do kilkudziesięciu centymetrów na minutę. Odbywa się on dzięki transpiracji, parciu korzeniowemu oraz właściowściom tkanki przewodzącej.
Transport w obrębie tkanek liścia przebiega poziomo, podobnie jak w korzeniu kanałami symplastycznym i apoplastycznym. Polega na przemieszczaniu się substancji od wiązek przewodzących przez komórki miękiszu do aparatów szparkowych, gdzie kończy się wyparowaniem wody z powierzchni liścia. Części nieużytecznej wody i soli mineralnych może być usunięta na zewnątrz liścia m.in. na skutek gutacji.

Najważniejszą rolę w regulacji ilości wody w roslinie odgrywa transpiracja, czyli utrata wody poprzez parowanie z zadziemnych części rośliny. Wpływa ona na pobieranie i transport wody z solami mineralnymi, umożliwia wymianę gazową, a także chroni roślinę przez przegrzaniem. Znane są trzy rodzaje transpiracji: kutykularna, szparkow i przetchlinkowa.
Transpiracja kutykularna zachodzi wprost przez zewnętrzną powierzchnię liścia, czyli przez skórkę pokrytą kutykulą. Jej intensywność zależy przede wszystkim od grubości warstwy kutykuli. Kutykula nie przepuszcza bowiem wody, ale może ją wchlaniać i dlatego pęcznieje. Jeśli ilość wody w kutykuli będzie odpowiednio duża, to zacznie ona wyparowywać z powierzchni rosliny. Ocenia się że tylko 1-3% wody paruje przez kutykulę. Rośliny żyjące w środowisku wilgotny, maja cienką kutykulę, a transpiracja kutykularna odgrywa u nich ogromną rolę. U rolslin środowisk suchych, zwykle o grubej kutykuli, ten rodzaj transpiracju praktycznie nie wystepuje.
U roslin więksozść wody wyparowyje zwykle w procesie transpiracji szparkowej. Jej intensywność jest zmienna i zalezy od wielu czynników. Czynniki wewnętrzne są związane z budową rosliny, m.in. z wiekością systemu korzeniowego oraz wielkością i strukturą anatomiczną aparatów szparkowych. Czynnikami zewnętrznymi to m.in. temperatura, światło, wilgotność powietrza, dostępność wody glebowej.
Wzrost temperatury zwieksza intensywność transpiracji z dwóch powodów: wpływa na mechanizmy otwierania aparatów szparkowych oraz zmniejsza wilgotność wilgotność względną powietrza. Podobnie jak temperatura na wilgotność powietrza działa wiatr, który usuwa wilgotne powietrze z bliskoego otoczenia liści. Im mniejsza wilgotność powietrza, tym większa jest różnica potencjału między rosliną a powietrzem i tym intensywnuejsza jest transpiracja.
Rola światła jako czynnika modyfikującego intensywność transpiracji wynika przede wszsytkom z jego wpływu na otwieranie aparatów szparkowych oraz ogrzewnaie blaszki liścia, Na intensywność transpiracju wpływa również dostepność wody glebowej, Jej niedobór powoduje zmniejszenie zawartości wody w tkankach liści, a to z kolei prowadzi do zamykania aparatów szparkowych.
Transpiracja przetchlinkowa odbywa się przez przetchlinki korka pokrywającego łodygi roślin drzewiastych. Różni się ona od transpiracji szparkowej tym, że przetchlinki nie zmniejszają swojej szerokości, co uniemozliwia regulację intensywności tego rodzaju transpiracji.

Roślina powinna pobierać taka ilość wody, aby móc równoważyć jej zużycie na własne potrzeby z utratą wody w wyniku transpiracji. Na tym polega zrównoważony bilans wodny, zachodzący w warunkach optymalnych dla rośliny. Bilans wodny może być dodatni, kirdy ilość pobieranej wody przewyższa jej straty (np. zwiędnięte rośliny uzupełniające deficyt wody), lub ujemny, kiedy straty wody są większe od jej pobieranej ilości.
W miarę pogłębiania się deficytu wody liście i niezdrewniałe lodygi wiodczeją, co określa się mianem więdnięcia. Może ono być przejściowe - wtedy powrót do normalnego stanu następuje zwykle w nocy, ponieważ o tej porze zmniejsza się transpiracja. W wypadku więdnięcia trwałego przywrócenie tugoru następuje wyłącznie wtedy, gdy zostanie dostarczona odpowiednio duża ilość wody. Jeśli do tego nie dojdzie, zachodzi więdnięcie nioedwracalne, kończące się śmiercią rołsiny.
Deficyt wody hamuje wiele procesów, m.in. fotosyntezę. Dlatego niedobór wody ogranicza lub nawet uniemozliwia np. kiełkowanie nasion czy wzrost organów. Zahamowaniu ulagają także procesy przewodzenia soli mineralnych oraz produktów fotosyntezy. Okres życia rośliny, w którym jest ona najbardziej wrażliwa na niedobór wody nazywa się okresem krytycznym. Dla wiekszości roślin dwuliściennych jest nim faza kwitnienia, a np dla uprawnych roślin jednoliściennych (m.in. zbóż) faza strzelana w źdźbło (wydłużania się łodygi po wytworzeniu pierwszego międzywęźla) i kłoszenia, czyli wytwarzania kwiatostanów.
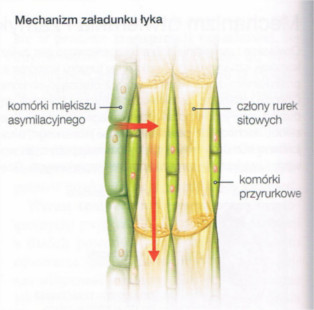
W procesie fotosyntezy w liściach roślin są produkowane węglowodany (głównie sacharoza) będące substancjami odżywczymi (asymilacyjnymi). Muszą być one dostarczone do wszystkich komórek. Transport asymilatów odbywa się w elementach przewdodzących łyka w górę i w dół rośliny. Istotną rolę odgrywają tu dwa zależne od siebie procesy: załadunek i rozładunek łyka.
Załadunek łyka odbywa się w organach odpowiedzialnych za wytwarzanie asymilatów, czyli głównie w liściach. Komórki miększu asymilacyjnego liści dostarczające sacharozę nazywa się donorami. Z donorów sacharoza przemieszcza się najpiwer do komórek przyrurkowych, a następnie do członów rurek sitowych przez liczne plazmodesmy. Transport ten wymaga nakładu energii, ponieważ odbywa się wbrew różnicy stężeń - stężenie sacharozy w komórkach towarzyszących (przyrurkowych jest do 20 razy większe niż w donorach.
Proces odwrotny, czyli rozładunek łyka, zachodzi w organach, które nie wytwarzają asymilatów lub wytwarzają je w niewystarczających ilościach. Następnie w ich odbierane sacharozy z elementów przewodzących łyka. Proces ten również wymaga nakładu energii. Komórki, które są miejscem przeznaczenia asymolatów, nazywa się akceptorami.
Mechanizmami transportu asymilatów wzdłuż elementów przewodzących łyka nie jest jeszcze w pełni wyjaśniony. Według jednej z hipotez transport ten odbywa się zgodnie z różnicą tugoru między miejszce w którym następuje załadunek łyka (liście), a miejscem w którym następuje jego rozładunek (np. korzenie). W czasie załadunku łyka we wnętrzu rurek sitowych następuje zwiekszenie stężenia sacharozy i obniżenie potencjału wody. To pociąga za sobą osmotyczny napływ wody z naczyń do rurek sitowych. Wytwarza się przez to ciśnienie hydrostatyczne , które tłoczy roztwór cukru w rurce sitowej.